На тему «Мхи» в школьной программе отведено 2
часа. Объем информации на заданную тему в данной
статье значительно превышает тот минимум,
который учитель может выдать ученикам за 2
академических часа. Тем не менее, автор все-таки
предлагает ее вам, дорогие учителя. С одной
стороны, это, конечно, программа
университетского курса, причем составленная
мировыми светилами бриологии. Но, с другой
стороны, автор думает, что хороший учитель – это
такой учитель, который знает больше, чем говорит.
Потому что в школьную программу не уместить
всего. Козьма Прутков по этому поводу в свое
время заметил: «Еще раз скажу – нельзя объять
необъятное». Следуя этому дельному совету, автор
все-таки о многом умолчал, изложив курс бриологии
в конспективной форме и снабдив его
многочисленными иллюстрациями, дабы облегчить
восприятие...
СИСТЕМАТИКА
Отдел Моховидные (Bryophyta).
I. Класс Листостебельные мхи, или
Мхи, или Бриопсиды (Bryopsida, или Musci).
Подклассы
1. Sphagnidae. 2. Bryopsidae.
3. Andreidae.
II. Класс Печеночники, или
Печеночные мхи (Marchantiopsida, или Hepaticopsida).
Подклассы
1. Jungermanniidae. 2. Marchantiidae.
III. Класс Антоцеротовые, или
Антоцеротопсиды (Anthocerotopsida).

Рис. 1. Представители основных групп
мохообразных: а – Sphagnidae; б – Bryopsidae; в – Andreidae; г
–
Jungermanniidae; д – Marchantiidae; е – Anthocerotopsida.

Рис. 2. Формы роста мхов: а – наземные, б
– эпифитные, в – водные, г – полуводные
Мхи – очень маленькие растения и
обычно растут там, где влажно. В сырых лесах и
безлесных местообитаниях они образуют сплошные
ковры. Но есть и виды, способные расти в крайне
сухих местообитаниях.
Так же, как и другие растения, мхи :
– являются многоклеточными
организмами, способными размножаться половым
путем;
– имеют специализированный орган, внутри
которого развивается зигота;
– имеют споры и спороносные органы, покрытые
кутикулой.
В отличие от других растений, мхи:
– не имеют лигнина в клеточных
стенках;
– сильно зависят от влажности воздуха (временно
прекращают жизнедеятельность в засушливый
период);
– не имеют настоящих корней.
ХАРАКТЕРИСТИКА ОСНОВНЫХ ГРУПП
БРИОФИТОВ
Класс Бриопсиды (Bryopsida)
Подкласс сфагновые мхи (Sphagniidae)
Размеры стеблей мхов измеряются в сантиметрах,
их листьев – в миллиметрах, а клеток – в
микрометрах.
Морфология

Рис. 3. Растения сфагнов, объединенные в
дернинку
|

Рис. 4. Головки сфагновых мхов
|
Стебель у сфагнов
прямостоячий, но отдельные растения не способны
расти прямо из-за отсутствия механической ткани.
Поэтому сфагнум всегда растет дернинками (рис. 3),
в которых отдельные растения опираются друг на
друга веточками, растущими из стебля
горизонтально, подобно тому, как могут опираться,
обнявшись, друг другу на плечи люди. На верхушке
стебля находится более или менее компактная
головка, состоящая из скученных вокруг точки
роста коротких молодых веточек (рис. 4). По мере
роста стебля вверх веточки вырастают в длину и
рассредоточиваются по стеблю отдельными пучками,
состоящими из горизонтально отстоящих и свисающих
вдоль стебля веточек (рис. 5). Отстоящими
веточками соседние растения в дернинке
сцепляются между собой. По пористым клеткам
листьев этих веточек осуществляется
горизонтальный транспорт веществ в сфагновой
дернинке. По клеткам листьев свисающих ветвей
осуществляется транспорт веществ по вертикали
(из почвенного раствора). Листостебельное
растение сфагна не имеет ризоидов.

Рис. 5. Пучок веточек у сфагна
Анатомия
Гаметофор сфагнов состоит из двух
типов клеток – живых, хлорофиллоносных, и
мертвых, лишенных содержимого крупных водоносных,
или гиалиновых, клеток (рис. 6). Стенки
последних укреплены спиральными волокнами,
предохраняющими их от разрыва при потере
тургора. У многих видов в стенках гиалиновых
клеток имеются поры, облегчающие всасывание воды
(рис. 6, 3). Так как листья, произрастающие на
веточке, налегают друг на друга, как черепица на
крыше дома (их так и называют – черепитчато-налегающими),
поры клеток соседних листьев накладываются друг
на друга, так что в олиственной веточке
образуется капиллярная система. Она
продолжается между веточками и соседними
растениями. Толщина листьев сфагновых мхов –
всего лишь в один слой клеток (рис. 7).

Рис. 6. Клетки сфагна: 1 – водоносная
клетка; 2 – хлорофиллоносная клетка; 3 – пора; 4 –
спиральные волокна
|

Рис. 7. Поперечный срез листа сфагна
|
Более сложное строение
обнаруживает стебель (рис. 8). В нем клетки, хотя и
дифференцированны, но не образуют
специализированных тканей, как у сосудистых
растений. Внутренняя часть стебля заполнена
тонкостенными клетками, осуществляющими как
транспорт веществ к точке роста, так и запасающую
функцию. Совокупность этих клеток называется сердцевиной
стебля. Сердцевина окружена узкими
толстостенными клетками склеродермиса,
выполняющего функцию коры и частично –
механическую функцию. Кнаружи от склеродермиса
имеется слой мертвых тонкостенных гиалиновых
клеток, образующих гиалодермис. В
гиалодермисе накапливаются и сохраняются запасы
влаги, осуществляется траспорт воды к точке
роста.

Рис. 8. Поперечный срез стебля сфагна: 1
– гиалодермис; 2 – склеродермис; 3 – сердцевина
|

Рис. 9. Спорогон сфагна
|
Спорогон сфагнов, как и у других мхов,
состоит из ножки и коробочки (рис. 9), но в отличие
от класса Bryopsida, ножка спорогона очень
короткая (ее функцию выполняет ложноножка,
образующаяся из гаплоидных клеток брюшка
архегония), а коробочка не имеет перистома.

Рис. 10. Стеблевой (1) и веточные (2)
листья сфагна
Листостебельное растение (гаметофор)
состоит из стебля с веточками или без них. Как на
стебле, так и на веточках у бриофитов имеются
листья, которые называются соотвественно стеблевыми
и веточными (рис. 10). Листья образуются в
результате деления в точке роста на верхушке
стебля. Вслед за листьями вырастают вначале
коротенькие веточки, которые, удлиняясь, как бы
«прихватывают» с собой в качестве «багажа» ранее
образовавшиеся листья. По мере роста апикальной
меристемы вверх, обособляется стебель с
веточками. Листья, которые попадают на стебель,
приобретают другую анатомию, в отличие от
веточных, строение которых не изменяется. У
молодых растений и растений, развивающихся в
неблагоприятных условиях, часто стеблевые
листья не отличаются от веточных. Такие растения
называют равнолистными, или изофильными.
В неблагоприятных условиях стеблевые листья
могут развиваться лишь наполовину. В этом случае
растения называют полуравнолистными, или гемиизофильными.
Нормально развитые растения называют неравнолистными,
или анизофильными (рис. 11).

Рис. 11. Онтогенез гаметофита у сфагнов:
1 – изофильные растения; 2 – гемиизофильное
растение; 3 – анизофильное растение
Экологические особенности и
распространение
Мхи способны заселять наиболее бедные
и влажные местообитания. В умеренной зоне и в
Арктике они образуют обширные торфяные болота.
Торф – это неразложившиеся спрессованные
остатки сфагновых мхов, которые сохраняются
практически в неизменном виде столетиями и
тысячелетиями. Многие болота таежной зоны имеют
возраст от 8 000 до 10 000 лет. Вместе со сфагнами
гнению не подвергаются и остатки других болотных
растений. Происходит это потому, что торфяные мхи
выделяют в окружающую среду целый букет
органических кислот (яблочную, лимонную,
уксусную, муравьиную и др.). В такой кислой среде
из организмов, питающихся растительным опадом,
способны существовать только некоторые виды
грибов. Но и их деятельность подавляется очень
сильным фунгицидом – сфагнолом (гликозидом с
фенольной основой, химическая формула которого
до сих пор точно не известна), который тоже
выделяют в почвенный раствор сфагновые мхи. Он же
оказывает и гербицидное действие на всходы
древесных растений. Поэтому на старых торфяных
болотах часто нет леса.
Но сфагны способны расти не только в
местообитаниях с бедным минеральным питанием.
Часто их можно встретить и в поймах лесных речек,
и на озерах, и у родников (выходах ключевых вод). В
таких местообитаниях они, правда, не образуют
сплошных ковров, а также торфа, так как
биохимический состав растений тех видов мхов,
которые здесь произрастают, не столь химически
агрессивен.
В подобных местообитаниях сфагны
произрастают преимущественно в тропической и
субтропической зонах, за исключением торфяных
болот Бразилии и Новой Зеландии. Собственно,
субтропики являются родиной сфагновых мхов, что
видно как по наличию в этой зоне примитивных
форм, так и по разнообразию видов. Так, во всем
мире насчитывается около 300 видов рода Sphagnum,
из которых в умеренной зоне Евразии и Америки
произрастает только около 50 (рис. 12). Несмотря на
более упрощенное строение, по сравнению с
представителями класса Bryopsida, сфагны
эволюционно более молодые.

Рис. 12. Ареал сфагновых мхов
Подкласс Настоящие мхи (Bryopsidae )
Морфология
В нижней части стебля у бриевых мхов
имеются нити из лишенных хлорофилла клеток – ризоиды,
которые выполняют функцию закрепления на
субстрате и проведения воды к стеблю (рис. 13).
Ризоиды образуются на ранней стадии прорастания
споры, параллельно с протонемой. Протонема
может быть нитчатой (похожей на водоросли) и
пластинчатой (рис. 14). Протонема с почками
возобновления листостебельных растений (поздняя
стадия развития протонемы) называется каулонемой
(рис. 15).
 Рис.
13. Гаметофит мха с ризоидами
|
 Рис. 14. Пластинчатая (1) и нитчатая
(2) протонемы мхов
|

Рис. 15. Каулонема (протонема с почками)
Анатомия
Стебель имеет относительно сложное строение
(рис. 16, 17). Внешний слой образует эпидермис, из
клеток которого могут вырастать многоклеточные
нити – ризоиды. Следующий за эпидермисом слой
называется склеродермис. Он состоит из узких
и длинных толстостенных клеток, выполняющих
проводящую и механическую функции. Самый
внутренний слой клеток называется центральным
пучком, он состоит из сосудов, очень
напоминающих сосуды других высших растений.
Различают два типа клеток сосудов – лептоиды
(похожие на клетки флоэмы) и гидроиды
(подобные клеткам ксилемы). Совокупность
лептоидов в стебле называют лептом, а
гидроидов – гидром (рис. 18). Помимо лептоидов
и гидроидов в центральном пучке имеются стереиды,
сходные с волокнами склеренхимы сосудистых
растений. В месте прикрепления стеблевого листа
к стеблю от центрального цилиндра ответвляется
пучок лептоидов и гидроидов, образующих листовую
вставку (аналогичную листовому следу у
сосудистых). Анатомия стебля сильно варьирует у
разных представителей бриофитов, у конкретных
видов в состав стебля могут входить не все
перечисленные компоненты или они могут
по-разному комбинироваться.
 Рис.
16. Схематическое строение стебля
Bryidae
|

Рис. 17. Поперечный срез стебля
кукушкина льна: 1 – лист; 2 – центральный
пучок; 3 – эпидермис
|

Рис. 18. Трехмерная диаграмма строения
стебля мха
|

Рис. 19. Транспорт воды гидроидами: 1 –
центральный пучок; 2 – листовая вставка
|
 Если
сравнивать строение ножки спорофита со стеблем,
можно обнаружить, что в состав и того и другого
входят гидроиды. Анатомия гидроидов очень сходна
с анатомией трахеид сосудистых: их клетки имеют
толстые стенки и удлиненные острые окончания.
Однако у гидроидов, в отличие от трахеид,
отсутствуют спиральные целлюлозные волокна, а их
стенки лишены лигнина. Гидроиды осуществляют
проводящую функцию (рис. 19) Если
сравнивать строение ножки спорофита со стеблем,
можно обнаружить, что в состав и того и другого
входят гидроиды. Анатомия гидроидов очень сходна
с анатомией трахеид сосудистых: их клетки имеют
толстые стенки и удлиненные острые окончания.
Однако у гидроидов, в отличие от трахеид,
отсутствуют спиральные целлюлозные волокна, а их
стенки лишены лигнина. Гидроиды осуществляют
проводящую функцию (рис. 19)
Строение лептоидов сходно со строением
ситовидных трубок сосудистых.
Лист большинства видов бриофитов имеют «жилку»,
строение которой сходно со строением стебля (рис.
20).
Рис. 20. Лист Bryopsidae с центральной жилкой
Жизненный цикл
Из гаплоидной споры вырастает
первичный росток – протонема (рис. 14), которая
развивается в протонему с почками – каулонему
(рис. 15). Со временем из почки вырастает тоже
гаплоидное листостебельное растение – гаметофор.
Вначале оно молодое, недоразвитое, но вскоре
набирает силы и приобретает привычный облик.
Совокупность гаметофора, протонемы и каулонемы
называется гаметофитом, или гаплоидным
поколением мха. У гаметофора мхов есть одна
примечательная особенность. Выражаясь образным
языком, он имеет вечно живущую часть – точку
роста (рис. 21), срок жизни которой не ограничен
(даже зимой), и временно живущую часть, или
«медленно разлагающийся труп», – все остальное
листостебельное растение, срок жизни которого
определен сезоном вегетации. На гаметофоре со
временем образуются гаметангии – антеридии
(мужские половые органы) (рис. 22, 1) и архегонии
(рис. 23, 2) (женские половые органы). Антеридии
– это «мешочки на ножках» со стенкой из одного
слоя клеток. Внутри антеридиев созревают антерозоиды
– мужские половые клетки, способные к
самостоятельному передвижению в водной среде.
Антеридии вырастают в пазухах веточных листьев,
которые отличаются по своему строению от
стерильных, имеют красноватую окраску и
называются антеридиальными (рис. 22, 2).
Совокупность антеридиев и покрывающих их
листьев на веточке называют перигонием.
Архегонии вырастают на стебле, как боковые
веточки. Сверху они прикрыты перихециальными
листьями (рис. 23, 1). Совокупность архегония и
покрывающих его листьев называют перихецием.
Архегонии устроены более сложно (рис. 23, 2). По
форме они напоминают колбочку. В нижней (широкой)
части архегонии имеют двухслойную стенку. Эта
его часть называется брюшком. Внутри брюшка
находится несколько яйцеклеток. Узкая часть
архегония – «горлышко колбочки» – имеет
одноклеточную стенку. Эта часть называется шейкой.
Антеридии и архегонии у видов сырых
местообитаний образуются на гаметофоре ранней
весной, сразу после схода снега, пока талая вода,
необходимая для передвижения антерозоидов, не
сошла. В шейке зрелого архегония образуется
слизистый канал, по которому антерозоиды
попадают в брюшко. В брюшке оплодотворяются
сразу несколько яйцеклеток, но в рост идет только
одна из них. Она дает начало спорофиту –
спороносному поколению мха с диплоидным набором
клеток. Спорофит состоит из коробочкиножке,
накрытой сверху, как плащом, колпачком –
остатками наружных стенок архегония. Коробочка
на ножке без колпачка (который часто отсутствует)
называется спорогоном (рис. 24). (в
зрелом состоянии – коричневого цвета) на

Рис. 21. Апикальная точка роста мха
(показана стрелкой)
|

Рис. 23. Архегоний и перихеций:
1
– архегоний; 2 – перихециальные листья; 3
– шейка архегония; 4 – яйцеклетка
|
 Рис.
22. Антеридиальная веточка мха: 1 – антеридии; 2 –
антеридиальные листья
|

Рис. 24. Спорогон: 1 – ножка; 2 –
коробочка; 3 – колпачок; 4 – перистом у открытой
коробочки; 5 – крышечка
Спорофит вырастает примерно через
месяц–другой после оплодотворения. Внутри
незрелой коробочки содержится спорогенная
ткань, состоящая из гаплоидных клеток. Зрелым
спорогон становится после мейотического деления
клеток спорогенной ткани, в результате которого
образуются гаплоидные споры.
Вегетативное размножение
Половое размножение у мхов изучено
достаточно хорошо. Однако существует множество
видов, которые вообще никогда не производят
спорогонов. Вегетативный способ размножения
характерен для всех видов мхов без исключения.
Новое листостебельное растение может развиться
буквально из любого фрагмента гаметофита, даже
из единственной клетки, а также (у некоторых
видов) из клеток ножки спорофита.
Кроме этого имеются и
специализированные органы для вегетативного
размножения. Это выводковые почки и тельца
(геммы) (рис. 25).

Рис. 25. Выводковые тельца мхов (геммы):
внизу – прорастающее выводковое тельце с
ризоидом
В зависимости от того, в какой части
гаметофита образуется спорофит, листостебельные
мхи делят на верхоплодные и бокоплодные.
Верхоплодные мхи имеют прямостоячий гаметофор,
как правило, без боковых веточек. Спорофит у них
вырастает из верхушки стебля (по крайней мере,
верхней его части). Бокоплодные мхи имеют
многочисленные боковые веточки, главный стебель
не прямостоячий, а полегающий. Спорогоны
образуются в латеральных частях стебля или на
верхушках боковых веточек (рис. 26).

Рис. 26. Верхоплодные (1) и бокоплодные (2)
мхи
Подкласс андреевые мхи (Andreidae )
Андреевые мхи – это группа наскальных
мхов (рис. 1, в). Они способны поселяться на
монолитной каменистой породе, буквально
вгрызаясь в каждую трещинку ризоидами. Формируют
довольно плотные дерновинки. В клеточных стенках
листьев андреевых мхов находится красный
пигмент, благодаря чему растения имеют
темно-красный цвет. Этот пигмент выполняет
функцию светофильтра для хлоропластов, которые у
этих мхов не способны нормально фунционировать
при полном освещении.
В мире насчитывается около 100 видов
андреевых мхов, большинство из которых
произрастают в полярных регионах северного и
южного полушарий. Некоторые виды растут в
альпийском поясе гор тропиков.
 Andreidae по некоторым признакам сходны
со сфагновыми мхами, по некоторым – с
печеночными и бриевыми. Так, их листостебельный
гаметофит по внешнему виду напоминает гаметофит
кукушкина льна (род Polytrichum). Так же, как и у
бриевых мхов, листья имеют центральную жилку. Так
же, как сфагновые мхи, андреевые имеют
пластинчатую протонему. Коробочка андреевых
мхов раскрывается на несколько створок (рис. 27),
как у печеночников, но в отличие от последних,
верхние концы створок остаются соединенными
друг с другом, что придает созревшему спорогону
вид японского фонарика. Так же, как и у сфагновых
мхов, у андреевых ножка спорогона очень
укорочена, так что коробочка выносится вверх с
помощью ложноножки, образующейся из гаплоидных
тканей архегония. Andreidae по некоторым признакам сходны
со сфагновыми мхами, по некоторым – с
печеночными и бриевыми. Так, их листостебельный
гаметофит по внешнему виду напоминает гаметофит
кукушкина льна (род Polytrichum). Так же, как и у
бриевых мхов, листья имеют центральную жилку. Так
же, как сфагновые мхи, андреевые имеют
пластинчатую протонему. Коробочка андреевых
мхов раскрывается на несколько створок (рис. 27),
как у печеночников, но в отличие от последних,
верхние концы створок остаются соединенными
друг с другом, что придает созревшему спорогону
вид японского фонарика. Так же, как и у сфагновых
мхов, у андреевых ножка спорогона очень
укорочена, так что коробочка выносится вверх с
помощью ложноножки, образующейся из гаплоидных
тканей архегония.
Рис. 27. Растение андреевого мха с
раскрытой коробочкой
Класс печеночные мхи (Hepaticae )
Название «печеночники» эти мхи
получили из-за внешнего сходства наиболее
распространенного в Европе их представителя –
маршанции – с формой печени. В Средние века часто
лекарственные свойства растений определяли по
внешнему сходству с соответствующим органом
человека. Считалось, что Бог пометил каждое
растение соответствующей формой, указывающей на
их свойства исцелять болезни тех органов, на
которые они похожи по внешнему виду.
В мире насчитывается около 8500 видов
печеночников, которые распространены во всех
климатических зонах – от тропиков до полярных
широт. Хотя среди них имеются виды, способные
расти в крайне засушливых местообитаниях или в
водоемах, большинство все же встречается в
умеренно увлажненных, таких как сырые луга,
берега речек.
Преобладающим поколением в жизненном
цикле печеночников является гаметофит, который
может быть как олиственным (подкласс
Юнгерманниевые), так и талломным (подкласс
Маршанциевые). Размеры его у разных видов
колеблются от 0,15 до 25 см (рис. 28, 29).

Рис. 28. Представитель подкласса
маршанциевых печеночников
|

Рис. 29. Представитель подкласса
юнгерманниевых печеночников
|
У юнгерманниевых печеночников
листья располагаются на стебле в двух боковых
рядах и, в отличие от бриевых мхов, не имеют жилки.
Кроме того, на так называемой «брюшной» стороне
стебля (т.е. там, где он соприкасается с землей) у
них имеются брюшные листья, или амфигастрии (рис.
30). Как юнгерманниевые, так и маршанциевые
прикрепляются к субстрату ризоидами.
Вегетативное размножение
печеночников осуществляется путем ветвления
стебля (вновь образовавшаяся веточка отделяется
от материнского растения и продолжает рост
самостоятельно) или с помощью
специализированных выводковых органов.
Последние представляют собой корзиночки с
выводковыми почками (рис. 31). По внешнему виду эти
корзиночки могут быть серповидными, чашевидными
или бутылковидными
.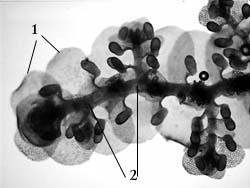
Рис. 30. Юнгерманниевый печеночник: 1 –
листья; 2 – амфигастрии

Рис. 31. Корзиночки с выводковыми
почками у маршанции
|

Рис. 32. Антеридии у маршанции
|
Генеративное размножение
осуществляется специализированными половыми
органами – антеридиями и архегониями (рис. 32).
Оплодотворение осуществляется в жидкой среде. Из
оплодотворенной яйцеклетки вырастает спорофит,
состоящий из коробочки со спорами на ножке (рис.
33, 1)

Рис. 33. Спорофит печеночников: 1 –
незрелый; 2 и 3 – зрелый, после рассеивания спор
Зрелая коробочка раскрывается на
четыре створки, благодаря чему происходит
рассеивание спор (рис. 33, 2, 3). Спора дает
начало новому гаметофиту.
Класс антоцеротовые (Anthocerotopsida)
Антоцеротовые – самый маленький класс в отделе
Bryophyta. Виды этого класса распространены по
всему земному шару и произрастают во влажных
тенистых местообитаниях. Гаметофит внешне похож
на таллом маршанциевых. Спорофит имеет
апикальную меристему и поэтому постоянно растет
вверх. Говорят, что он напоминает рога (рис. 34).
Споры созревают, начиная с нижней его части.
Зрелый спорофит раскрывается сразу по всей
длине.
|

