К отделу плауновидных (Lycopodiophyta)
относятся наиболее древние из высших растений, встречающихся
сейчас на нашей планете. Правда, в современной
флоре плауновидные представлены небольшим
числом родов и видов, которые не играют
существенной роли в формировании растительного
покрова – своего расцвета эти растения
достигали около 300 млн лет назад – в конце
палеозойской эры.
Все современные представители
плауновидных, а их около 1000 видов, многолетние
травянистые растения, часто вечнозеленые. Побеги
большинства растений этой группы покрыты
спирально расположенными мелкими листьями – микрофиллами.
Характерно для плауновидных также
дихотомическое или вильчатое ветвление побегов.
Подземные части этих растений обычно
представлены корневищем с придаточными корнями
и видоизмененными листьями. У некоторых видов
подземные части побегов образуют своеобразный
орган, несущий расположенные по спирали корни и
называемый ризофором (от греч. rhiza –
корень и phoros – несущий), или корненосцем.
Надземные и подземные побеги плауновидных
нарастают ограниченное время, поскольку со
временем клетки верхушечных меристем теряют
способность к делению.
Спороносные листочки (спорофиллы)
плауновидных по форме, размерам и цвету могут
быть похожи на вегетативные листья (трофофиллы)
или в той или иной степени отличаться от них.
Чередуясь со стерильными листьями, спорофиллы
образуют на протяжении стебля спороносные зоны
или собраны в расположенные на концах ветвей
компактные образования – стробилы. У
некоторых ископаемых форм плауновидных
спорофиллы сидели на стебле вперемежку с
вегетативными листьями, не образуя ни
спороносных зон, ни стробилов.
Среди плауновидных есть равно- и
разноспоровые растения, гаметофиты (заростки)
которых, соответственно, либо обоеполые (т.е. на
них образуются как мужские, так и женские половые
клетки), либо однополые. Есть между ними и другие
отличия. Заростки современных равноспоровых
(представителей класса плауновых) созревают в
течение 1–15 лет, достигая в длину от 2 до 20 мм. При
этом они развиваются в подземных или
полуподземных условиях, и питаются в
значительной степени или исключительно за счет
образования союза с почвенными грибами. Заростки
разноспоровых плауновидных (к ним относятся все
представители класса полушниковых) развиваются
обычно в течение нескольких недель. Они вообще
лишены хлорофилла и живут за счет питательных
веществ, содержащихся в споре. Поэтому они очень
мелкие и даже в зрелом виде лишь слегка выступают
за пределы оболочки споры.
Половые органы, развивающиеся на
заростках плауновидных, представлены антеридиями,
в которых развиваются сперматозоиды с двумя или
многими жгутиками, и архегониями, в которых
развиваются яйцеклетки. Для того чтобы
сперматозоид мог достичь яйцеклетки, необходимо
наличие капельно-жидкой воды.
Надо заметить, что для того чтобы
наблюдать эту фазу в цикле развития плаунов, как
и других высших споровых растений, нужно
проявить наблюдательность и терпение
натуралиста-исследователя – разыскать
крошечные заростки под травяно-кустарничковым
ярусом леса очень непросто.
Оплодотворенная яйцеклетка у
плауновидных сразу же, не впадая в состояние
покоя, развивается в растение нового бесполого
поколения – спорофит (рис. 1).
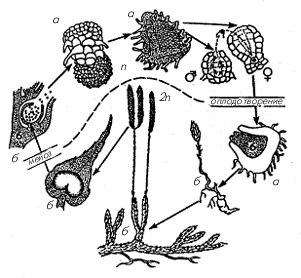
Рис. 1. Чередование поколений в
жизненном цикле плауна: а – гаметофит; б –
спорофит
В настоящее время на нашей планете
встречаются представители двух классов отдела
плауновидных – плауновых и полушниковых, или
шильниковых.
Плауновые (Lycopodiolsida) представлены
в современной флоре единственным одноименным
порядком (Lycopodiales) и семейством. Все плауновые
– вечнозеленые наземные или эпифитные (т.е.
растущие на стволах и ветвях деревьев) растения с
прямостоячими, полегающими, свисающими,
ползучими или лазящими побегами. Высота
прямостоячих побегов у наземных растений или
свисающих (у эпифитов) обычно не превышает 1–1,5 м,
но длина ползучих побегов может достигать 10 м и
более.
Эпифитные формы плаунов характерны
для тропических лесов. В нашей же стране плауны
можно встретить в зеленомошных хвойных лесах.
Это представители родов плаун (рис. 2–4),
например, плаун годичный (Lycopodium annotinum) и плаун
булавовидный (L.clavatum) и баранец – баранец
обыкновенный (Huperzia selago).
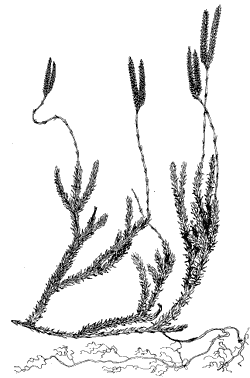
Рис. 2. Плаун булавовидный (Lycopodium clavatum)

Рис. 3. Плаун годичный (Lycopodium annotium)
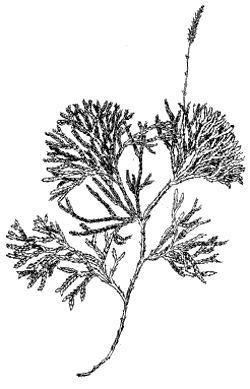
Рис. 4. Плаун сплюснутый (Lycopodium complanatum)
У первых двух названных видов стебли
ветвятся неравнодихотомически, образуя ползучий
стебель (главную ось) и отходящие от него боковые
полегающие побеги с ограниченным ростом.
Спороносные листья у этих плаунов собраны в
верхушечные стробилы — колоски.
У баранцов стебли ветвятся
равнодихотомически – в побегах этих растений
нельзя выделить главную ось и боковые
ответвления. Кроме того, у баранцов, в отличие от
собственно плаунов, спорофиллы не отличаются от
обычных ассимиляционных листьев и не образуют
выраженных спороносных колосков.
Класс полушниковых (Isoetopsida) в
современной флоре представлен двумя порядками –
селагинелловыми (Selaginellales) и полушниковыми (Isoetales).
К селягинеллам относится около 700
видов растений (рис. 5–6), единственного рода
селягинелла (Selaginella). Многие селягинеллы
внешне похожи на мхи. Их побеги, как и у плаунов,
характеризуются дихотомическим ветвлением,
ограничены в росте и обильно ветвятся.

Рис. 5. Селагинелла: а – сибирская; б –
крючковатая; в – завертывающаяся; г – Мартенса; д
– тамарисколистная
Большинство селягинелл – тене- и
влаголюбивые растения тропических лесов, где они
зачастую образуют на почве сплошной ковер. У
немногочисленных ксерофильных представителей
этого рода, обитающих в степях или на открытых
скалах, побеги при высыхании улиткообразно
скручиваются, окружая центральную точку роста
сферическим покровом. Один из видов этой
экологической группы селягинелл – селягинелла
тамарисколистная (S.tamariscina) – нередко
встречается на Дальнем Востоке. На сухих
открытых местах Сибири обычны селягинеллы
скальная (S.rupestris) и кровавопятнистая (S.sanguinolenta).
По всей территории нашей страны встречается и
более влаголюбивый вид, характерный для сырых
зеленомошных прогалин хвойных лесов и сырых
лугов, — селягинелла обыкновенная (S. selaginoides)
(рис. 6). Однако это маленькое (не более 10 см в
высоту) растение обычно остается незамеченным,
теряясь среди зеленых мхов. Да и в целом в наших
условиях селягинеллы не играют существенной
роли в растительных сообществах.
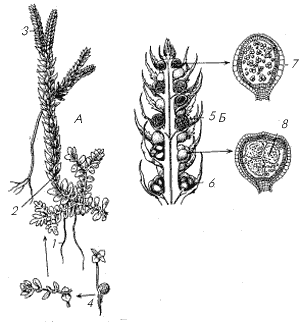
Рис. 6. Селагинелла обыкновенная: А –
спорофит: 1 – придаточный корень, 2 – стебель с
листьями, 3 – стробил, 4 – молодой спорофит; Б –
стробил со спорангиями: 5 – микроспорангий, 6 –
мегаспорангий, 7 – микроспора, 8 – мегаспора
Селагинеллы, как и все представители
класса полушниковых, – разноспоровые растения. У
большинства из них микро- и мегаспорофиллы
находятся в одном стробиле (рис. 6,Б), а иногда
спорофиллы сходны с остальными листьями побега и
стробил не выражен. Заростки селягинелл сильно
редуцированы и обычно развиваются, не покидая
оболочек рассеившихся спор. В этом случае из
микроспоры выходят уже сформированные
сперматозоиды. Даже после оплодотворения
зародыш спорофита долгое время продолжает
использовать для роста запасные питательные
вещества мегаспоры.
Порядок полушниковых также
включает единственный род Isoetes1,
представленный примерно 70 видами растений. Почти
все полушники – погруженноводные (очень редко
наземные) травы с коротким вертикальным
утолщенным стеблем и розеткой
линейно-шиловидных цилиндрических листьев на
верхушке. Основание стебля несет двулопастное
утолщение (ризофор), на котором образуются корни.
Стебель и ризофор полушников имеют
рудиментарный камбий и способны к
незначительному вторичному утолщению –
фактически эти растения с их вертикальными
неветвящимися стволиками являются современными
карликовыми формами гигантских древовидных
лепидодендронов, в изобилии населявших нашу
планету в палеозойскую эру.
Очень крупные мега- и микроспорангии
полушников образуются в ямках, лежащих в
основании листьев на их верхней поверхности. В
мегаспорангиях образуется от 50 до 2350 мегаспор, а
число микроспор в микроспорангии может
достигать миллиона.
Распространены полушники очень
широко, встречаясь почти во всех районах земного
шара от тропиков до Арктики. Наиболее типичное
место их обитания – озера с чистой и прозрачной
водой. Встречаются они и по мелководьям морей. В
России можно встретить полушники озерный (I.lacustris,
рис. 7), щетинистый (I.setacea), азиатский (I.asiatica),
морской (I.maritima) и др. Из-за
антропогенного загрязнения водоемов все эти
виды в настоящее время довольно быстро вымирают.

Рис. 7. Полушник озерный (Isoetes lacustris)
Ископаемые плауновидные
Наиболее древней и примитивной
группой плауновидных является порядок
астероксиловых (класс плауновых),
представители которого известны из отложений
среднего и верхнего девона. Астероксилон
(рис. 8) был травянистым растением с четким
разделением на подземную и надземную части, а
также на стебель и мелкие шиловидные листья,
которые густо покрывали стебли, как у
современных плаунов. Обилие воздухоносных
полостей в стебле свидетельствует о земноводном
(полупогруженном) образе жизни растения. Стебли
астероксилона отходили от горизонтально
распростертой дихотомически ветвящейся
подземной части. В ее коре, как и в коре стебля,
были обнаружены гифы и споры грибов, с которыми,
по-видимому, астероксилон находился в симбиозе.

Рис. 8. Астероксилон (Asteroxylon)
В девонских отложениях найдены также
остатки растений из другого, целиком вымершего в
палеозойскую эру, порядка класса плауновых – протолепидодендронов
(рис. 9). Обладая характерными особенностями
плауновидных, эти растения отличались от других
представителей отдела вильчато-разветвленными
листьями.

Рис. 9. Протолепидодендрон Шари
(Protolepidodendron schariatum)
Во второй половине палеозойской эры
плауновидные достигли максимального расцвета:
представители многочисленных родов и семейств
были широко распространены по земному шару и
занимали различные экологические ниши. Но
основную роль в формировании растительного
покрова в то время играли представители класса
полушниковых. Среди них особенно выделялись
своими размерами лепидодендроны, или чешуедревы
(Lepidodendron) (рис. 10) из порядка
лепидодендровых. Лепидодендроны имели
колонновидный главный стебель (ствол) высотой до
30 м и диаметром более 1 м у основания. Большая
часть ствола была образована паренхимной тканью;
древесина же занимала небольшой объем. Крона
формировалась в результате многочисленных
последовательных дихотомических ветвлений
стебля. Ствол молодого растения был густо покрыт
длинными шиловидными (до 1 м длиной при ширине
около 1 см) листьями. По мере роста растения
листья постепенно опадали и на их месте
оставались листовые рубцы с выходами
воздухоносной ткани, через которые происходил
газообмен растения с окружающей средой. По
межклетникам этой ткани, пронизывающей все
органы растения, воздух проходил и в корневую
систему, находящуюся в болотистой почве. На
подземных осях корневой системы (ризофорах)
располагались по спирали корни длиной до 50 см и
более. Листья лепидодендронов имели на нижней
стороне две устьичные ложбинки, на дне которых
располагались многочисленные устьица. При таком
погруженном положении устьиц капли воды,
конденсируемые на поверхности растения из
атмосферной влаги, не могли закупоривать их и
испарение воды (которое во влажной атмосфере
тропических болот и так шло с небольшой
скоростью) не прерывалось. Это имело большое
значение для поддержания восходящего тока воды в
гигантском стволе и громадной кроне.
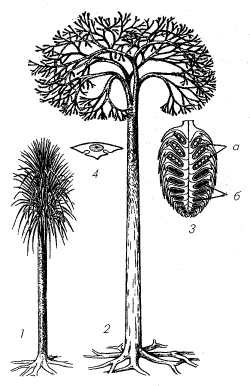
Рис. 10. Лепидодендрон (Lepidodendron). 1 –
молодое растение; 2 – взрослое растение; 3 –
стробил: а – мегаспорангии, б – микроспорангии; 4
– схема поперечного среза листа
Стробилы лепидодендронов,
располагавшиеся на концах ветвей, могли
достигать в длину 50 см и диаметра 5 см (рис. 10, б).
В каменноугольных лесах кроме древовидных
представителей полушниковых произрастали и
травянистые формы плауновых и селагинелловых.
Лепидодендроны и отчасти сигиллярии вместе с
рядом гигантских хвощевидных образовали на
Земле основные запасы каменного угля.

Рис. 11. Сигиллярия (Sigillaria)
 В В
пермское время на смену лепидодендронам пришли
более низкорослые и менее разветвленные сигиллярии,
или печатницы (Sigillaria, рис. 11), которых в
мезозое сменили плевромейи (Pleuromeia)
– невысокие (1–2 м), неразветвленные растения с
одним верхушечным стробилом (рис. 12). В некоторых
районах Земли плевромейи образовывали по
берегам морей заросли, возможно, напоминающие
современные мангры. Одновременно с уменьшением
величины и степени разветвленности верхней
части оси полушниковых уменьшались размеры и
число ветвей ризофора. Современный полушник
условно можно считать лепидодендроном,
созревающим в первый год жизни. Гигантские же
древовидные формы плауновидных вымерли еще до
конца палеозойской эры, т.е. не позже 250 млн лет
назад.
Рис. 12. Плевромейя Штенберга (Pleuromeia
stenbergii)
«Ведьмины круги»
У некоторых плаунов, обладающих
ползучими побегами (например, булавовидного,
заливаемого, сплюснутого, растущих на бедных
песчаных почвах в редкостойных сосняках, на
вересковых пустошах или по берегам озер и
окраинам болот в умеренной зоне), отмечено
образование «ведьминых кругов», или колец. Эти
кольца, появление которых легенды связывают с
местами шабаша нечистой силы, представляют собой
группы растений, возникшие при вегетативном
размножении спорофитов, выросших на одном или
нескольких расположенных рядом заростках.
Побеги молодых растений, ветвясь и нарастая в
длину, сначала образуют небольшую куртину,
которая постепенно приобретает округлое,
овальное или вееровидное очертание.
Дальнейший центробежный рост главных побегов и
ветвей и отмирание их в основании и приводят к
образованию замкнутых или незамкнутых полос –
«ведьминых колец», ширина которых зависит от
скорости роста и продолжительности жизни
ползучих побегов.
У плауна баранца диаметр куртины в благоприятных
для роста условиях (влажные хвойные леса)
достигает иногда 2 м, а возраст таких куртин
исчисляется десятилетиями, а иногда и
столетиями.
У плауна сплюснутого некоторые кольца имеют
диаметр 40 м и более – таких размеров они, как
показывают расчеты, могут достигнуть за 150–300
лет.
Хозяйственное значение плаунов
Позвоночные травоядные животные не
едят плауны – некоторые виды этих растений
ядовиты и содержат сильный парализующий яд,
сходный по характеру действия с ядом кураре. В то
же время их издавна использовали в народной
медицине и ветеринарии, входят плауны и в
современную фармакопею разных стран. Их
используют при лечении алкоголизма, никотинизма,
глазных болезней. (Не следует, конечно,
заниматься самолечением: это может привести к
сильному отравлению вплоть до смертельного
исхода.)
Использовали плауны и как источник зеленой,
желтой, синей красок.
Богатые жирными маслами споры некоторых плаунов
раньше применяли в пиротехнике (при изготовлении
взрывчатых веществ и для получения световых
эффектов), а ныне их продолжают использовать при
фасонном литье – при соприкосновении с жидким
металлом споры, покрывающие поверхность формы,
вспыхивают, и образовавшийся газ способствует
получению гладкой поверхности детали.
Традиционно используются споры плаунов в
медицине в качестве порошка (ликоподия) для
присыпки (при пролежнях; как детская присыпка), а
также для обсыпки пилюль. Но в целях сохранения
запасов плаунов рекомендуют при заготовках
отрезать лишь спороносные колоски, стараясь не
повредить другие части растения. И уж конечно,
нельзя использовать плауны для изготовления
гирлянд и украшения помещений: слишком медленно
они растут, чтобы так расточительно их
уничтожать.
Селагинеллы не имеют большого хозяйственного
значения – некоторые их виды с красивой ажурной
листвой иногда выращивают как теневыносливые
декоративные растения. А некоторые виды
полушников разводят в аквариумах.
Какие растения называют высшими?
В группу высших растений (некоторые
ботаники придают ей ранг подцарства) объединяют
наиболее эволюционно продвинутые формы
растений. Их тело обычно расчленено на стебель,
корень и лист (по этой причине эту группу
называют еще листостебельными, или побеговыми,
– Cormophyta, или Cormobionta). Зигота у них, в
отличие от низших растений, дает начало
многоклеточному зародышу, который затем
развивается в растение, производящее споры, –
спорофит (на этом основании их называют еще зародышевыми
– Embryophyta, Embryobiontia). В отличие от
водорослей, высшие растения хорошо
приспособлены к жизни в наземно-воздушной среде:
их органы состоят из дифференцированных тканей
– проводящих, покровных, механических, основных,
меристематических и др. Высшие растения делят на
две группы – высшие споровые и семенные. К
семенным относятся отделы голо- и
покрытосеменных растений, размножающихся
семенами, а к высшим споровым – отделы риниевых,
зостерофилловых, моховидных, плауновидных,
псилотовидных, хвощевидных, папоротниковидных. У
представителей этих семи отделов (три из которых
известны только в ископаемом виде) процессы
споро- и гаметогенеза разобщены во времени и
пространстве: растения, образующие споры
(спорофиты), и растения, образующие гаметы,
являются самостоятельными организмами, ведущими
раздельное существование. У семенных растений
споро- и гаметогенез тесно сопряжены:
редуцированный женский гаметофит развивается
прямо на спорофите, а мужской переносится к
яйцеклетке целиком в виде пыльцевого зерна.
|

