Одной из приоритетных задач современной биологии, несомненно, является создание обобщенной теории эволюции экосистем.
При этом у экологов может возникнуть сильный соблазн начать ее строить в
одиночку, опираясь исключительно на данные по динамике современных
сообществ и игнорируя «измышления» палеонтологов. А почему бы и нет?
Есть тому пример: эволюцию всех конкретных таксонов животных и растений
реконструировали, как всем известно, при самом минимальном участии
палеонтологов, используя в основном лишь данные сравнительной анатомии.
Для этого достаточно сходные рецентные организмы располагают в
виде последовательностей, трактуемых как отражение реальной
преемственности в ряду «предок — потомок» (например, тупайя — лемур —
низшая обезьяна — человекообразная обезьяна — человек), а уже потом эти филетические ряды дополняют ископаемыми членами (в нашем случае — австралопитеком, питекантропом и неандертальцем).
Хотя злые языки из числа антиэволюционистов и
утверждают, будто филетические ряды мало чем отличаются от известной
карикатуры XIX века, изображающей «Чудесное превращение короля
Луи-Филиппа в грушу», это, разумеется, не так. Настоящая проблема в
другом: при реконструкции исторических последовательностей косвенный, «сравнительно-анатомический», метод действительно предпочтителен (поскольку метод прямой, «палеонтологический», имеет крупный встроенный дефект в виде принципиальной
неполноты летописи), однако он годится не для всякого типа
биологических объектов. Этот метод приложим лишь к тем объектам, которые
хотя бы в принципе обладают филогенезом, т.е. историческим развитием; иными словами, система должна обладать хорошей памятью относительно своих предшествующих исторических состояний.
В случае таксона память эта обеспечивается
передаваемым от предка к потомку «текстом» в виде нуклеотидной
последовательности. А вот экосистема подобного наследственного субстрата
лишена; она обладает лишь элементами онтогенеза, т.е. индивидуального
развития (экогенетические сукцессии, о которых речь пойдет в главе 12),
но филогенеза у нее, судя по всему, не бывает в принципе. Так что для
изучения истории экосистем «сравнительно-анатомический метод» не
годится, и палеонтология, как это ни парадоксально, неизбежно будет
играть здесь существенно большую роль, чем при реконструкции филогенезов
отдельных таксонов.
Итак, создавать теорию эволюции экосистем должны, на
паритетных началах, экологи и палеонтологи, причем первым, по идее,
следует создавать основу для актуалистической интерпретации тех фактов,
что накоплены вторыми. К сожалению, в процессе такого «заочного
сотрудничества» между экологами и палеонтологами часто возникают
недоразумения, связанные с неверным пониманием границ применимости
каждого из используемых ими методов. Одну из таких «конфликтных
ситуаций» можно разобрать на примере взаимоотношений хищника и жертвы в
докембрийских планктонных экосистемах).
В современной экологии существует достаточно хорошо обоснованная (в том числе и математическими моделями) концепция оптимального питания хищника. Согласно ей, консумент стремится выбирать пищевые объекты таким образом, чтобы разность между энергосодержанием пищи и суммарными энергозатратами на ее добывание оказывалась максимальной. Для каждого конкретного хищника существует определенный размерный оптимум жертв,
с наибольшей частотой поедаемых им (лев питается в основном антилопами,
хотя в составе его добычи могут оказаться и буйвол, и кролик). В свою
очередь, у жертвы имеется целый ряд эволюционных стратегий, защищающих ее популяции от выедания (увеличение скорости размножения, приобретение криптической окраски или ядовитости и т.д.); экспериментально показано, что одной из таких стратегий является уход за пределы размерного оптимума.
Из наблюдений над сезонными изменениями в популяциях планктонных ракообразных
и коловраток было установлено, что это действительно так; при этом
жертва по-разному защищается от тех хищников, которые лишь немного
превосходят ее по величине, и тех, которые относятся к следующему
размерному классу. Защитой от врагов первого типа (выедающих особей
среднего и мелкого размера) является увеличение собственых
размеров жертвы за пределы оптимума выедания. Против врагов второго
типа (выедающих самых крупных из жертв) такая стратегия бесполезна,
здесь надо уменьшать свои размеры. Защитой же от комбинированного воздействия хищников обоих типов может служить изменение формы тела — образование шипов и прозрачных выростов
кутикулы (их периодическое появление в популяциях дафний — хорошо
известный факт). Это позволяет увеличить реальные размеры (создав помехи
для врагов первого типа), сохраняя при этом возможно меньшую заметность
для врагов второго типа. Основываясь на этой экологической концепции, а
также на эмпирических данных о распространении в геологической истории
основных размерных классов фитопланктона (до 16 мкм, 16–60 мкм, 60–200
мкм, свыше 200 мкм) и его морфологических групп (оболочка гладкая или с
выростами), М. Б. Бурзин в 1987 году реконструировал основные этапы
формирования планктонной экосистемы (рис. 22).
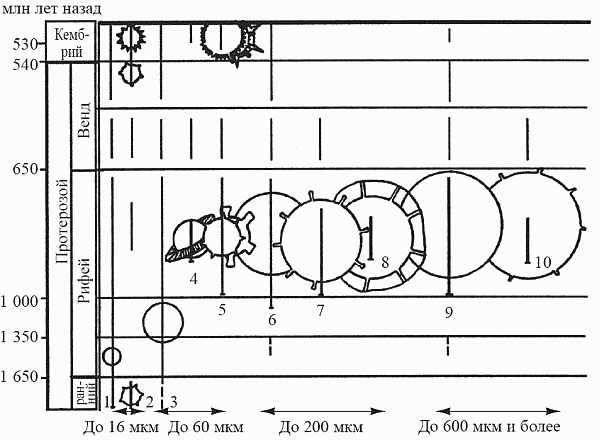
Рис. 22. Схема распространения в позднем
докембрии и раннем кембрии основных размерных и морфологических групп
фитопланктона (по Бурзину, 1987)
До начала рифея (1,65 млрд лет назад) фитопланктон
состоял почти исключительно из малоразмерных форм (до 16 мкм). Можно
предполагать, что консументы в это время либо были представлены самым
мелким размерным классом (назовем его условно «инфузория»), либо
отсутствовали вовсе. Впрочем, первое более вероятно: из этого времени
известны и мелкие шипастые формы, что может быть первым в истории
появлением защитных структур у фитопланктона в ответ на пресс выедания
первым зоопланктоном. Для эукариотного фитопланктона, обладающего
способностью разрешать «противоречие Виленкина» за счет активных
передвижений, выгодно сохранять минимальные размеры тела
(что улучшает соотношение «поверхность — объем»). Поэтому появление в
раннем рифее (1,6 млрд лет назад) форм из следующего размерного класса
(16–60 мкм) может рассматриваться как попытка фитопланктона отразить
усиление пресса хищников класса «инфузория»: уйти за размерный оптимум
выедания путем увеличения размера.
Эта стратегия использовалась фитопланктоном на
протяжении всего рифея, т.е. почти миллиард лет. В середине рифея
(1,4–1,0 млрд лет назад) появляются акритархи размерного класса 60–200
мкм; это может рассматриваться как реакция на появление консументов
следующего размерного класса — «коловратка». К концу же позднего рифея
(700 млн лет назад) фитопланктонные организмы достигают максимального
размера за всю историю: помимо доминирующей размерной группы 200–600 мкм
в это время появляются, например, гигантские акритархи диаметром до 2–3
мм — это физиологический предел размера для одноклеточных.[21] Увеличение размера как главная защитная стратегия
(при явно подчиненном значении выростов оболочки) свидетельствует о
сильном прессе зоопланктона класса «коловратка» (при отсутствии более
крупных хищников). Вообще тенденция к однонаправленному изменению
размера для жертвы имеет смысл лишь при существовании единственного размерного класса хищников — в противном случае тут же попадешь «из огня да в пòлымя».
А вот дальше произошел вендский фитопланктонный
кризис: разнообразие акритарх, достигнув незадолго до появления
эдиакарской фауны (600 млн лет назад) своего максимума, резко падает,
причем крупноразмерные формы исчезают полностью. Попытки объяснить этот
кризис тем, что фитопланктонные сообщества могли столкнуться в это время
с глобальным дефицитом CO2 при избытке O2 (такое
сочетание действительно угнетает развитие микроводорослей), кажутся
довольно наивными. Гораздо убедительнее смотрится гипотеза Бурзина о
том, что кризис возник в результате появления консументов следующего
размерного класса — «циклоп» (до 2 мм), составляющих основную массу
современного зоопланктона. От этого врага уже не могла спасти ставшая
стандартной стратегия опережающего ухода в следующий размерный класс,
ибо фитопланктон уже «уперся» в физиологический предел размеров тела.
Лишь к началу кембрия фитопланктону удалось дать адекватный эволюционный
ответ на комбинированное воздействие консументов разных классов: с
этого момента (и доныне) доминирующей защитной стратегией для
фитопланктоных организмов становится образование шипов, выростов,
экваториальной каймы и т.п.
Мы уделили столько внимания этой гипотезе по той
причине, что рисуемые ею взаимоотношения хищника и жертвы кажутся резко
противоречащими современной экологической парадигме, основанной на так называемых циклических балансовых моделях (типа модели Лотки — Вольтерры). Биомассы предыдущего и последующего трофических уровней соотносятся примерно как 10:1 (принцип трофической пирамиды).
Ясно, что именно популяция хищника должна подчиняться изменениям в
популяции жертвы, но никак не наоборот. Отсюда, как кажется,
естественным образом следует и так называемая филогенетическая реактивность хищника:
жертва увеличивает скорость передвижения — хищник вынужден перейти к
охоте из засады; жертва одевается панцирем — хищник начинает создавать
инструменты для его взлома; именно в таком порядке.
Однако многие палеонтологи (В. А. Красилов,
В. В. Жерихин, А. С. Раутиан) приходят в последнее время к выводу о том,
что балансовые модели — это путь к познанию функционирования сообществ, но не их эволюции.
Все необратимые эволюционные процессы (от глобальных экосистемных
кризисов до филогенеза отдельных таксонов) протекают не благодаря, а
вопреки сохранению экологического равновесия. Функционирование сообществ
основано на циклических процессах, протекающих с отрицательной обратной
связью; для того же, чтобы началось развитие, она должна разрушиться и
смениться на положительную обратную связь.
Применительно к системе «хищник — жертва» это должно
выглядеть так. Любой эволюционный успех жертвы (увеличение скорости
передвижения и т.п.) немедленно становится для хищника ощутимым;
эволюционные приобретения хищника вообще-то безразличны жертве,
поскольку в норме тот все равно изымает лишь небольшую часть ее
популяции. До тех пор, пока хищник остается в таком реактивном
состоянии, система пребывает в равновесии. Но как только эффективность
хищника превысит некое пороговое значение, жертва начинает «замечать»
хищника, поскольку теперь он подрывает ее ресурсы. В тот самый момент,
когда реактивной становится жертва, а не хищник, в системе и возникает
положительная обратная связь по типу «гонки вооружений». Система, выйдя
из равновесия, начинает эволюционировать в некоем (совсем не обязательно «правильном») направлении.
На эволюционную роль верхних трофических уровней
экосистемы можно посмотреть и с иной точки зрения. Их взаимодействия с
нижними уровнями являются не столько энергетическими (описываемыми в
терминах трофической пирамиды), сколько информационными. Здесь может
быть использована аналогия с постиндустриальным обществом, в
котором решающее значение приобретает не уровень производства энергии
(как это было в индустриальную эпоху), а оптимизация управления
энергопотоками за счет информационных связей. Именно таким «блоком
управления» экосистемы и являются консументы, деятельность которых
вызывает эволюционные изменения у их жертв. Как заметил А. С. Раутиан,
само выражение «эволюционные стратегии хищника и жертвы» не вполне
правильно: у хищника стратегия есть, а вот у жертвы — одна только тактика.
О степени же автономности этого «блока управления»
дают наглядное представление именно те палеонтологически
документированные ситуации, когда он производит чересчур
эффективного хищника, который, по выражению А. Г. Пономаренко, «проедает
экосистему насквозь» (как это и произошло при возникновении
зоопланктона размерного класса «циклоп»). Любой эколог скажет не
задумываясь, что «этого не может быть, потому что не может быть
никогда!», и будет совершенно прав — но прав именно в рамках своего,
экологического, масштаба времени. В эволюционном же масштабе времени
такая ситуация (ее можно условно назвать «изобретением абсолютного
оружия») не просто возможна — она, как мы увидим далее, служила реальным
механизмом запуска целого ряда крупнейших экосистемных перестроек и
филогенетических революций.
|

